
فهرست مطالب
- 1 داستان
- 2 پایه
- 2.1 Vererbung
- 2.2 تنوع ژنتیکی
- 2.3 همسانی
- 2.4 قیاس
- 3 عامل تکاملی
- 3.1 جهش
- 3.2 ترکیب مجدد
- 3.3 انتخاب
- 3.4 Gendrift
- 4 توسعه پیچیدگی بیولوژیکی
- 5 بازسازی فرآیند تکامل
- 5.1 تاریخچه فسیلی
- 5.2 سیستماتیک فیلوژنتیک
- 5.3 جغرافیای زیستی مقایسه ای
- 5.4 رشد جنینی مقایسه ای
- 6 ادراک عمومی
- 7 ادبیات
- 8 پیوندهای وب
- 9 اثبات فردی
داستان
صفات موجودات زنده به شکل ژنهایی کپی میشوند رمزگذاری میشوند که در طی تولید مثل و به فرزندان منتقل میشوند. منجر جهش به انواع مختلف ( الل ) این ژن ها می شود که می تواند منجر به پیدایش ویژگی های تغییر یافته یا جدید شود. این گونه ها و نوترکیبی ها منجر به تفاوت های ارثی در قالب تنوع ژنتیکی بین افراد می شود. تکامل زمانی اتفاق میافتد که فراوانی آللهای خاص در یک جمعیت ( فرکانس آلل در مخزن ژنی ) تغییر میکند و صفات مربوطه را نادرتر یا رایجتر در جمعیت میکند. این امر یا از طریق انتخاب طبیعی (نرخ های متفاوت بقا و تولیدمثل بر اساس این صفات)، انتخاب جنسی یا به طور تصادفی از طریق رانش ژنتیکی رخ می دهد.


گسست قاطع با مفهوم قبلی طبقات یا انواع گونهشناختی ثابت در زیستشناسی با نظریه تکامل توسط انتخاب طبیعی که توسط چارلز داروین و آلفرد راسل والاس بر حسب تغییر جمعیت در طول زمان فرمولبندی شد، رخ داد. این اهمیت تغییرپذیری و فرآیندی که توسط انتخاب طبیعی در موجودات زنده ایجاد میشود، برای اولین بار توسط چارلز داروین در کتاب منشاء گونهها در سال 1859 به طور کامل و عمیق نشان داده شد . در حدود سال 1900 اصول وراثت شناخته شد (از آنجایی که تحقیقات قبلی گرگور مندل نادیده گرفته شده بود) و در حدود سال 1910 اهمیت کروموزوم ها. به نظر میرسید که این با مفاهیم داروینیسم که بر تغییر تأکید داشت و ژنتیک که انتقال ثابت صفات را آموزش میداد، در تضاد بود. تنها از دهه 1930 بود که فرآیندهای گزینش میتوانست با قوانین وراثت مندل مطابقت داده شود . نظریه ترکیبی تکامل توسعه یافت. او تکامل را به عنوان تغییر در طول زمان در بسامدهای نسبی آلل ها ( فرکانس های آللی ) در یک جمعیت تعریف کرد. این نظریه از طریق گزاره های توصیفی و علّی خود به اصل سازماندهی اصلی زیست شناسی مدرن تبدیل شد و توضیح مناسبی برای پیدایش تنوع حیات بر روی زمین ارائه کرد.
در سال 1944، کار اسوالد اوری و همکارانش شواهد قوی ارائه کرد که DNA حامل اطلاعات ژنتیکی است، زیرا قبلاً پروتئینها حاوی اطلاعات مربوطه بودند. همراه با رمزگشایی ساختار DNA توسط روزالیند فرانکلین ، جیمز واتسون و فرانسیس کریک در سال 1953، اساس فیزیکی وراثت روشن شد. از آن زمان، ژنتیک مولکولی نیز به بخش مرکزی زیست شناسی تکاملی تبدیل شده است . [3]
اصول اولیه
وراثت
با استفاده از نخود، گرگور مندل نشان داد که وراثت در واحدهای باریک تعریف شده (گسسته) رخ می دهد، به این صورت که صفات از نسل والدین به فرزندان به ارث می رسند، و این ویژگی ها گسسته هستند: اگر یکی از والدین نخودی گرد و دیگری نخود فرنگی چروکیده داشته باشد، آنگاه فرزندان به ارث می رسند. مخلوطی نیست بلکه نخودهای گرد یا چروکیده است. مندل همچنین نشان داد که صفات والدین توسط فرزندان به شیوه ای کاملاً تعریف شده و قابل پیش بینی، یعنی طبق قوانین مندلی به ارث می رسد . تحقیقات او مبنایی برای مفهوم صفات گسسته و ارثی، ژن ها بود. [4] کار مندل به این سوال دیرینه پاسخ داد که چرا گونه های صفت در جمعیت ها پایدار می مانند. در نگاهی به گذشته، باید متوجه شد که این یک تصادف بزرگ بود که او چیزی جز صفات مجزا را انتخاب نکرد، زیرا بسیاری از صفات دیگر (مثلاً با توجه به ارتفاع گیاه) تأثیرات ژنتیکی و همچنین محیطی پیچیدهتری داشتند.
تحقیقات بعدی اساس فیزیکی ژن ها را نشان داد و DNA را به عنوان ماده ژنتیکی شناسایی کرد. ژن ها به عنوان نواحی خاصی از DNA بازتعریف شده اند. DNA توسط موجودات زنده به عنوان کروموزوم ذخیره می شود. یک مکان خاص روی یک کروموزوم، جایگاه ژنی (یا به اختصار جایگاه ) و گونهای از یک توالی DNA در یک مکان خاص، آلل نامیده میشود . کپی برداری از DNA ناقص است و تغییرات ( جهش ) در ژن ها باعث ایجاد آلل های جدید می شود و بنابراین بر صفات کنترل شده توسط آن ژن ها تأثیر می گذارد. این رابطه ساده بین یک ژن و یک صفت در بسیاری از موارد وجود دارد، اما ویژگیهای پیچیده، مانند مقاومت در برابر بیماری، توسط ژنهای زیادی که با هم کار میکنند (“چند ژنی”) کنترل میشوند. [5]

تنوع ژنتیکی
تنوع یا تنوع ژنتیکی ناشی از جهش در DNA، جابجایی افراد بین جمعیت ها ( جریان ژن )، و اختلاط ژن ها در طول تولید مثل جنسی ( بازترکیب ) است. در برخی از اشکال حیات، مانند باکتری ها و گیاهان، تنوع نیز با اختلاط مواد ژنتیکی بین گونه ها از طریق جریان افقی ژن و هیبریداسیون ایجاد می شود. علیرغم همه این فرآیندهایی که باعث تغییرپذیری می شوند، بیشتر قسمت های DNA یک گونه ( ژنوم ) در همه افراد یک گونه یکسان است. [8] تغییرات نسبتاً کوچک در ژنوتیپ (بخشی از ژنوم که ویژگیها را کد میکند) میتواند تأثیر قابلتوجهی بر فنوتیپ (همه ویژگیهای ژنتیکی یک فرد، به عبارت ساده، بر ظاهر خارجی تعیینشده ژنتیکی) داشته باشد. به عنوان مثال، تفاوت در توالی های DNA بین شامپانزه ها و انسان ها تنها پنج درصد است.
فنوتیپ از تعامل ترکیب ژنتیکی فردی، ژنوتیپ آن، با محیط حاصل می شود. بنابراین تنوع صفات وراثتی در یک جمعیت منعکس کننده تنوع ژنوم در آن جمعیت است. فراوانی انواع صفات فردی می تواند در یک جمعیت متفاوت باشد و در رابطه با سایر آلل های ژن بزرگتر یا کوچکتر شود. همه نیروهای تکاملی با ترویج این تغییرات در بسامدهای آللی در یک جهت یا جهت دیگر عمل می کنند. تغییرپذیری یک صفت زمانی از بین میرود که یک آلل به فرکانس ثابتی برسد، یعنی زمانی که از جمعیت ناپدید میشود یا زمانی که همه آللهای موجود قبلی را جایگزین میکند.
همسانی
اندام های پستانداران مانند ستون فقرات، چشم ها، مجرای گوارشی، ریه ها مشابه سایر مهره داران است. این شباهت ها بر اساس نسب از یک جد مشترک است. این شباهت ها در زیست شناسی همسانی نامیده می شود. یافته های اسکلت فسیلی امکان تشخیص همسانی ها و در نتیجه به دست آوردن سرنخ هایی از یک اصل و نسب مشترک را فراهم می کند. در مسیر تکامل، بسیاری از موجودات زنده شیوه زندگی خود را تغییر داده اند. به دلیل تنوع ژنتیکی و انتخاب طبیعی، تغییر عملکردی اندام ها اتفاق افتاد. این منجر به انطباق ساختمان با عملکرد مربوطه شد.
مقایسه
اندام های مشابه دارای پلان های ساختاری اساسی متفاوتی هستند، اما شباهت هایی دارند که مستقل از اصل و نسب آنهاست. دلیل این امر سازگاری با عملکردهای برابر است. نمونه ای از این خال دست حفاری و بیل حفاری خال جیرجیرک است که بسیار شبیه هم هستند. در حالی که دست خال دارای اسکلت استخوانی است، جیرجیرک خال دارای اسکلت بیرونی کیتینی است. کریکت مول و مول دارای پلان های ساختاری متفاوتی از اندام های مشابه خارجی خود هستند. شباهت های مشابه از نظر فیلوژنتیکی مستقل از یکدیگر هستند، اما اجازه می دهد تا در مورد شرایط محیطی مشابه و روش های زندگی نتیجه گیری شود.
عوامل تکاملی
در زیست شناسی، عوامل تکاملی فرآیندهایی هستند که مخزن ژنی (مجموع همه تغییرات ژنی در یک جمعیت ) را تغییر می دهند. این در درجه اول از طریق تغییر در فراوانی آلل در مخزن ژن جمعیت رخ می دهد. این فرآیندها عامل اصلی تغییرات تکاملی هستند.
عوامل تکاملی ضروری که مخزن ژنی را تغییر میدهند (مجموع همه گونههای ژنی در یک جمعیت) جهش ، نوترکیب ، انتخاب و رانش ژن هستند.
جهش

جهش ها می توانند در هر نقطه ای از بدن خارج از خط زایا ، یعنی خارج از سلول های تولید مثلی رخ دهند و سپس جهش های سوماتیک نامیده می شوند. برخلاف جهشهای مولد، این جهشها هیچ تأثیر مستقیمی بر نسلهای بعدی ندارند، در صورتی که جهش سوماتیک تناسب اندام ارگانیسم ناقل را مختل کند و در نتیجه احتمال انتقال ژنهای خود از نظر آماری کاهش یابد. جهشهای زایا جهشهایی هستند که میتوانند توسط فرزندان از طریق خط زایا به ارث برسند . آنها مربوط به سلول های تخمک یا اسپرم و پیش سازهای آنها قبل و در حین اووژنز یا اسپرم زایی هستند. آنها معمولاً هیچ تأثیری بر ارگانیسم حاملی که در آن قرار می گیرند ندارند.
جهش ها و اثرات آنها را می توان شناسایی کرد: به عنوان مثال، توسعه آنزیم های جدید در میکروارگانیسم ها را می توان در حرکت سریع به دلیل زمان تولید کوتاهتر مشاهده کرد. های تازه ایجاد شده نمونه هایی از آنزیم هستند نایلونازها .
نوترکیبی
نوترکیبی ها بازآرایی ژن های موجود هستند. آنها می توانند به عنوان بخشی از نوترکیبی پاراجنسی احتمالاً از نظر فیلوژنتیکی قدیمی تر (در پروکاریوت ها و برخی قارچ ها) و همچنین به عنوان بخشی از تولید مثل جنسی رخ دهند. در دومی، که تقریباً برای همه گیاهان و جانوران معمول است، بین نوترکیبی داخل کروموزومی از طریق نوترکیب آللها در کروموزومها (در نتیجه تقاطع به مناسبت اولین تقسیم میوز) و نوترکیبی بین کروموزومی از طریق نوترکیب کل کروموزوم ها در مجموعه کروموزوم ها .
انتخاب
انتخاب زمانی اتفاق میافتد که افراد با ویژگیهای مطلوب برای بقا و تولید مثل بتوانند فرزندان بیشتری نسبت به افراد بدون آن صفات تولید کنند. به این ترتیب، سازگاری های بهبود یافته با شرایط محیطی می تواند در طول نسل ها برای جمعیت ایجاد شود. به عنوان بخشی از چنین تغییراتی در خصوصیات، یک گونه همچنین می تواند به عنوان بخشی از گونه زایی به گونه های جدید تقسیم شود . انتخاب “عادی” بین افراد گونه های مختلف صورت می گیرد و به آن انتخاب طبیعی نیز گفته می شود . یک مورد خاص “انتخاب مصنوعی” یا پرورش مورد استفاده توسط انسان است که به عنوان مثال مسئول بسیاری از نژادهای سگ است. به محض اینکه سگ های اهلی مجبور به زنده ماندن در طبیعت هستند، که یک پدیده گسترده در بسیاری از کشورهای جهان است، تنها ژنوتیپ های خاصی از طریق انتخاب طبیعی غالب می شوند. سگها از نظر اندازه، ویژگیهای رنگی و رفتار نسبتاً یکنواخت میشوند، زیرا ویژگیهای «افراطی» پرورشیافتهتر در محیط طبیعی مضر هستند و در نتیجه سگهای مربوطه موفقیت باروری کمتری دارند.

یک مورد خاص از انتخاب یا انتخاب طبیعی، انتخاب جنسی است که به صورت درون گونه ای (یعنی در یک گونه) عمل می کند: انتخاب برای ویژگی هایی که حضور آنها مستقیماً با موفقیت جفت گیری از طریق انتخاب ترجیحی شریک ارتباط دارد. [12] صفات تکامل یافته از طریق انتخاب جنسی به ویژه در نر حیوانات رایج است. اگرچه این صفات ممکن است شانس بقای افراد نر را کاهش دهد (به عنوان مثال، از طریق شاخ های مزاحم، تماس های جفت گیری یا رنگ های روشن)، چنین مردانی معمولاً موفقیت باروری بالاتری دارند. [13]
سیستم های کمکی و اجتماعی بودن موارد خاص دیگری را نشان می دهد: در بیش از 200 گونه پرنده و حدود 120 گونه پستاندار، ساختارهای اجتماعی را می توان یافت که در آن برخی از افراد، حداقل به طور موقت، بدون تولید مثل خودشان عمل می کنند و در عوض از همنوعان در تولید مثل خود حمایت می کنند. . این در تناقض آشکار با تزهای داروین است. با این حال، مطالعات روی این سیستمهای کمکی نشان داده است که هر چه ارتباط نزدیکتر یاریکنندگان با فرزندانی که قرار است بزرگ شوند، بیشتر باشد، درجه کمک بیشتر میشود. [14] از آنجایی که بخشی از ژنوم کمک کننده و فرزندان خارجی پرورش یافته یکسان است، مددکار می تواند بخشی از ژنوم خود را با وجود انکار تولیدمثل خود منتقل کند. از آنجایی که انتخاب دیگر بر اساس فنوتیپ نیست، بلکه بر اساس ژنوتیپ است، ریچارد داوکینز اصطلاح “ژن خودخواه” را برای این موارد و موارد مشابه ابداع کرد. [15] در مورد حشرات اجتماعی مانند مورچه ها اجتماعی و زنبورهای ، اکثریت ماده ها تا پایان عمر از تولید مثل خودداری می کنند. اجتماعی بودن با کاهش تناسب اندام تکاملی این زنان به دلیل یک ویژگی ژنتیکی مرتبط نیست. هاپلودیپلوئیدی ) با خواهرانشان بیشتر از فرزندان بالقوه خودشان مرتبط هستند. هنگام بزرگ کردن خواهران، آنها بخش بزرگی از ژنوم خود را نسبت به زمانی که دختران خود را بزرگ می کنند منتقل می کنند.
جندریفت
رانش ژنتیکی تغییر در فراوانی آلل ها از نسلی به نسل دیگر است که به این دلیل اتفاق می افتد که آلل های یک نسل از فرزندان از نظر آماری نمونه تصادفی از آلل های نسل والد هستند و بنابراین انتخاب آنها نیز مشمول خطای تصادفی است. حتی در صورت عدم انتخاب، فرکانس های آللی در طول زمان افزایش یا کاهش می یابد و در نهایت به 0% یا 100% می رسد (“تثبیت” آلل). بنابراین نوسانات فراوانی آلل در نسل های متوالی می تواند منجر به ناپدید شدن تک تک آلل ها از جمعیت به طور تصادفی شود. بنابراین دو جمعیت مجزا با فرکانسهای آللی در ابتدا برابر میتوانند از طریق نوسانات تصادفی به دو جمعیت مجزا با مجموعهای از آللهای متفاوت از هم جدا شوند. [17]
اینکه انتخاب طبیعی یا رانش ژنتیکی تأثیر بیشتری بر سرنوشت جهشهای جدید داشته باشد به اندازه جمعیت و قدرت انتخاب بستگی دارد. [18] انتخاب طبیعی در جمعیت های بزرگ غالب است و رانش ژنتیکی در جمعیت های کوچک. در نهایت، مدت زمانی که طول می کشد تا یک آلل به یک فرکانس ثابت در یک جمعیت از طریق رانش ژنی برسد (یعنی تا زمانی که 0٪ یا 100٪ افراد در جمعیت آلل را حمل کنند) به اندازه جمعیت بستگی دارد. این در جمعیت های کوچکتر سریعتر اتفاق می افتد. [19]
بنابراین اندازه یک جمعیت (به طور دقیق تر اندازه جمعیت مؤثر ) تأثیر عمده ای بر سیر تکامل دارد. به عنوان مثال، اگر جمعیتی از تنگنای ژنتیکی عبور کند (اندازه جمعیت به طور موقت بسیار کم)، همچنین بخش زیادی از تنوع ژنتیکی خود را از دست می دهد. جمعیت به طور کلی در حال همگن شدن است و بسیاری از انواع نادر را از دست می دهد. چنین «گلوگاههایی» میتواند ناشی از حوادث فاجعهبار، نوسانات آب و هوایی، مهاجرت یا تقسیم جمعیت و البته فشارهای انسانی باشد.
توسعه پیچیدگی بیولوژیکی
یک نتیجه مهم از فرآیند تکامل، توسعه پیچیدگی بیولوژیکی است. [20] تکامل ارگانیسم های بسیار پیچیده ای تولید کرده است. با این حال، تعریف یا اندازه گیری سطح عددی پیچیدگی در زیست شناسی بسیار دشوار است. ویژگی هایی مانند اندازه ژنوم، تعداد ژن، تعداد انواع سلول یا مورفولوژی به عنوان پارامترهای ممکن پیشنهاد شد.
معمولاً موجوداتی که نسبت به رقبای خود از نرخ تولید مثل بالاتری برخوردارند، از مزیت تکاملی برخوردارند. به منظور تولید مثل سریعتر و تولید فرزندان بیشتر، موجودات زنده می توانند به سمت سادگی بیشتر تکامل یابند، زیرا پس از آن به منابع کمتری برای تولید مثل نیاز دارند. یک مثال خوب انگل هایی مانند مالاریا پاتوژن Plasmodium و Mycoplasma هستند – این موجودات خواصی را که به دلیل انگلی زائد شده اند از دست می دهند.
یک اصل و نسب همچنین می تواند پیچیدگی خود را از دست بدهد، زمانی که یک ویژگی پیچیده خاص در محیط به سادگی مزیت انتخابی ایجاد نکند. حتی اگر از دست دادن این صفت مزیت انتخابی ایجاد نکند، در صورت خنثی بودن (که هیچ ضرر انتخابی فوری ایجاد نمی کند) این ویژگی می تواند از طریق تجمع جهش ها از بین برود.

اگر یک گرایش فعال به سمت پیچیدگی ( ارتوژنز ) در تکامل وجود داشت (همانطور که به طور گسترده در قرن 19 فرض می شد)، می توان انتظار داشت که رایج ترین مقدار ( حالت ) پیچیدگی در میان موجودات به طور فعال در طول زمان افزایش یابد.
با این حال، افزایش پیچیدگی را می توان با یک فرآیند غیرفعال نیز توضیح داد: با این فرض
- که پیچیدگی کاملاً تصادفی تغییر می کند (بدون ترجیح یک جهت خاص در معنای غایت شناختی) و
- که حداقل پیچیدگی وجود دارد
میانگین پیچیدگی بیوسفر در طول زمان افزایش می یابد. این شامل افزایش واریانس است، اما حالت تغییر نمی کند. سپس این تمایل برای برخی از موجودات زنده با پیچیدگی فزاینده در طول زمان وجود دارد، اما این بر درصد کمتر و کمتری از موجودات زنده تأثیر می گذارد. برخی از موجودات گهگاه به سمت راست تکامل می یابند، بنابراین “دم” سمت راست را در توزیع پیچیدگی طولانی تر می کنند . برخی نیز به سمت چپ حرکت می کنند، اما در نهایت با محدودیتی در سمت چپ منتهی می شوند که توسط حداقل الزامات زندگی تعیین شده است (برای مثال ، حداقل ژنوم را ببینید).
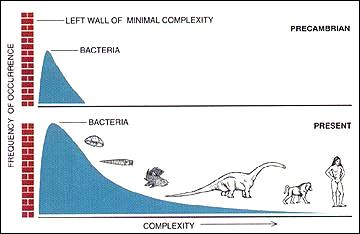
در حالت غیرفعال، هر شکل ظاهری تکامل ذاتاً (فعالانه/تلولوژیکی) که منجر به ارگانیسمهای پیچیدهتر میشود، ناشی از ادراک انسان است که بر معدود موجودات بزرگ و پیچیده در انتهای سمت راست توزیع پیچیدگی تمرکز میکند (خود انسان و سایر موجودات “بالاتر” حیوانات و گیاهان) و ارگانیسمهای سادهتر و بسیار رایجتر را نادیده بگیرید (نک . ماده تاریک میکروبی ) – البته به استثنای روزهای اولیه زمین، زمانی که فقط موجودات زنده با سازماندهی ساده وجود داشتند. مدل غیرفعال پیشبینی میکند که اکثر گونهها پروکاریوتهای میکروسکوپی هستند ، همانطور که توسط تخمینهای موجود [ 30 تا 106 109 گونههای پروکاریوت ] در مقایسه با تخمینهای تنوع 106 تا 3106 برای یوکاریوتها ( ارگانیسمهای سلولی پیچیده: پروتیست ها ، گیاهان ، قارچ ها و حیوانات – از جمله انسان ها) تایید شده است. پروکاریوتها ( باکتریها و آرکیاها ) تا به امروز رایجترین و موفقترین آنها باقی ماندهاند و این حالت موقعیت خود را در میان این موجودات تغییر نداده است.
استفن جی. گولد و همکاران. توزیعی که در ابتدا فقط برای موجودات سلولی کمی بیشتر در سمت چپ ادامه داد نشان داده شده بود را می توان با در نظر گرفتن ویروس ها . حداقل اندازه سازمانی (“دیوار چپ”) ویروس ها هنوز بسیار کوچکتر از موجودات سلولی است. با این حال، این با تسلیم استقلال از ارگانیسم های میزبان اتفاق می افتد . مهم است که تنوع مورد انتظار در این زمینه حتی بیشتر از مورد پروکاریوت ها باشد. حالت توزیع بیشتر به سمت چپ تغییر می کند. [33]
بازسازی فرآیند تکامل
فرآیندهای بازسازیپذیر تکامل زمینی – جهتگیریها و طبقهبندیهای زمانی – را میتوان بر اساس پیشینههای فسیلی و تحلیل الگوها و فرآیندهای اخیر بازسازی کرد. فرآیندهایی که در گذشته با توجه به جهت، ترتیب زمانی و سرعت تکامل رخ داده اند را می توان با اطمینان بیشتر بازسازی کرد، هر چه شواهد مستقل تر بتوانند به روشنگری رویدادهای تاریخی کمک کنند. اساساً فرض بر این است که سیر فرآیندهای زمین شناسی و بیولوژیکی در گذشته به روز است، یعنی از این فرض که فرآیندهای بیولوژیکی، اکولوژیکی و زمین شناسی در گذشته بر اساس اصول یکسان یا مشابه آن صورت می گرفته است. امروز مشاهده و اندازه گیری شود. برای این منظور، آزمایشهایی نیز میتوانند انجام شوند که تا حدی میتوانند بر روی فرآیندهای گذشته پیشبینی شوند.
در مورد اشکالی که فسیل کمی به جا گذاشته اند یا هیچ فسیلی از خود به جای گذاشته اند، که شامل تقریباً تمام پروکاریوت ها و اکثر های یوکاریوتی تک یاخته و همچنین همه موجودات غیراسکلتی دیگر مانند کرم ها ، چتر دریایی ، راب ها و غیره می شود، فقط مقایسه های اخیر انجام می شود. (امروزه) جانوران یا گیاهان با بررسی معقولیت انتقادی همزمان نتایج حاصل از آنها به کار گرفته می شوند. در اینجا، بازسازی فرآیندهای گذشته در درجه اول بر اساس شباهت مولکولی با استفاده از روش تجزیه و تحلیل روابط فیلوژنتیک انجام می شود. اصل ساعت مولکولی می تواند به نقاط اتصال دودمان های مختلف ( کلادها تعیین تاریخ تقریبی ) کمک کند. فرآیندهای تکامل مشترک ، برای مثال جذب سابق باکتری در سلولهای باستانی، که در آنجا (شاید دو میلیارد سال پیش، تخمین آن دشوار است) به میتوکندری و کلروپلاستها بهعنوان درون همزیستی وجود دارد . سلولهای یوکاریوتی را میتوان عملاً تنها از دادههای اخیر در مورد شباهتهای ژنتیکی مولکولی و همچنین شباهتهای متابولیسم و ساختار درونهمزیستها استنباط کرد .
اغلب اشکال دریایی اسکلتی به صورت فسیل یافت می شوند، گونه های بسیار کمیاب تر از سرزمین اصلی آب های داخلی (رودخانه ها، دریاچه ها). معمولاً فسیلهای بسیار کمی از مناطق کوهستانی، از باتلاقها و آبهای چشمه، و عموماً از مناطق خشک سابق روی زمین وجود دارد، زیرا به طور کلی امکان تعبیه و نگهداری در چنین مکانهایی وجود ندارد.
فسیل فسیلی

شوند، نشانههایی از چارچوب زمانی فرآیندهای تکاملی را ارائه میدهند فسیلها ، که میتوانند از نظر ریختشناسی بررسی ، اما بسیاری از خواص بیولوژیکی-اکولوژیکی، مانند زیستگاههای پرجمعیت، شیوههای حرکت یا حتی گاهی اوقات رفتار اجتماعی (مثلاً اگر زندگی میکنند ) از آنها ارائه میشود. در بسته یا ازدحام رخ می دهد) را می توان خواند. با توجه به توالی عمودی لایههای سنگدار فسیلی ، جوامع موجودات فسیلی را میتوان بر اساس (اصل چینهشناسی به ترتیب زمانی ) آورد. در حالی که این در ابتدا فقط اطلاعاتی در مورد “سن نسبی” ارائه می دهد (کدام فسیل ها زودتر بودند، کدام بعد؟)، با کمک روش های رادیومتری می توان مناسب ( ژئوکرونولوژی ) در سنگ ها و/یا فسیل های موجود در آن ها تاریخ گذاری مطلق انجام داد.
سن فسیلها اطلاعاتی را در مورد زمان طول تاریخ فیلوژنتیک گروههای فردی و موجودات زنده وقوع نوآوریها و رویدادهای شکاف (تابشهای تطبیقی) در ارائه میکند. به عنوان مثال، از یافتههای فسیلی با جدول دادهای مشخص شده است که (به استثنای چند مورد مانند کلودینا ) [34] همه گروههای جانوران اسکلتی برای اولین بار در یک پنجره زمانی باریک در اوایل تا اواسط دوره کامبرین ، حدود 540 تا 500 میلیون سال پیش ظاهر شدند. . [35] این که تا چه حد ناشی از نوآوریهای بیولوژیکی بود یا تا چه اندازه شرایط محیطی به گونهای تغییر کرد که تشکیل اسکلت و رسوبات از نظر شیمیایی و فیزیکی امکانپذیر شد، هنوز سؤالی است که در نهایت روشن نشده است.
اشکال انتقالی فسیلی (شکل های موزاییکی و حلقه های گمشده ) شواهد مستقلی از رابطه بین گروه های بزرگ سیستماتیک بر اساس مطالعات روی موجودات زنده هستند. نمونههای معروف عبارتند از « دایناسورهای پردار »، آرکئوپتریکس و پرندگان گروه Jehol بهعنوان اشکال انتقالی از آمنیوتهای غیرپرنده (بهطور عامیانه «خزندگان» یا – به طور کلاسیک – «دایناسورها»، که به درستی «دایناسورهای غیر پرنده» نامیده میشوند، هر دو پارافیلتیک واحد . ) و پرندگان مدرن [36] و همچنین پاندریکتس ، تیکتالیک و ایکتیوستگا به عنوان اشکال انتقالی بین ماهیان استخوانی و مهره داران زمینی . [37] در بهترین حالت، انتقال بین گروه اصلی و مشتق شده با توالی یافته های فسیلی که از نظر مورفولوژیکی با کاهش سن بیشتر شبیه به گروه جدید می شوند، اثبات می شود.
روندهای تکاملی در بسیاری از موارد به خوبی در گروه های سیستماتیک ثبت شده است، به عنوان مثال. به عنوان مثال در پستانداران اسبی انگشتی، به اندازه روباه و برگ خوار در اوایل دوره سوم های امروزی طی چندین مرحله میانی از شکل های چند : اسب تکامل یافتند .
در نهایت، اسناد ثبت فسیلی در تنوع گروه های سیستماتیک افزایش و کاهش می یابد. بریدگی های جانوران انقراض های دسته جمعی هستند که در آن تعداد گونه های فسیلی در دوره های زمانی کوتاه زمین شناسی به شدت کاهش یافت و برخی از گروه های بزرگ به طور کامل ناپدید شدند یا تنوع آنها بسیار کاهش یافت. شناختهشدهترین، هرچند نه بزرگترین، انقراض جانوران، انقراض گسترده « دایناسورها » (به استثنای گروه پرندگان) و سایر گروههای بزرگ در پایان دوره کرتاسه است. طاقچه های اکولوژیکی که آزاد شدند می توانند متعاقباً به عنوان بخشی از فرآیند تشعشع توسط پستانداران و پرندگان مدرن اشغال شوند.
مروری بر وقوع فیلاهای جانوری و گیاهی در زیر تاریخ تکامل ارائه شده است .
سیستماتیک فیلوژنتیک
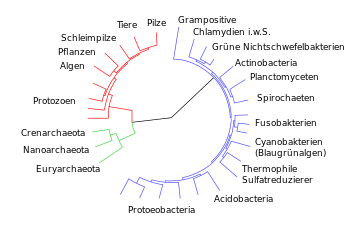
مقایسه ویژگیهای موجودات در چارچوب سیستماتیک بیولوژیکی نشان داد که ویژگیها در ترکیبهای دلخواه ظاهر نمیشوند، بلکه در سیستمی از شباهتهای درجهبندی شده ظاهر میشوند. گروههایی از ویژگیها را میتوان از یکدیگر متمایز کرد، که بر اساس آن موجودات اخیر را میتوان گروهبندی کرد ( تاکسون منفرد ، تاکسون )، خلاصه (طبقهبندی) و به صورت سلسله مراتبی مرتب کرد.
داروین این امکان را بهعنوان نشانهای قوی در نظر گرفت که همه موجودات زنده یک اصل و نسب مشترک دارند. برای اینکه این گروهبندیها (نظامسازیها) روابط واقعی را منعکس کنند ( سیستماتیک فیلوژنتیک ویژگیها به عنوان یک معیار مرکزی )، همسانی معرفی شد ، یعنی تشابه تنها زمانی معنادار است که ویژگیها یا اندامهای مربوطه را بتوان به همان ویژگیهای اصلی ردیابی کرد. . آفریقا که برخی از آنها به دلیل آبدار بودن شبیه کاکتوسهای آمریکا و خانواده اسپورگ به هم هستند، تشکیلات مشابهی از خود نشان میدهند. ظاهر اغلب مشابه آنها نشان دهنده رابطه نزدیکتر و تبار از یک اجداد مشترک نیست.
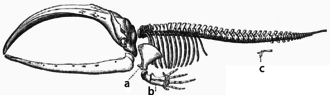
یک مورد خاص از صفات همولوگ، صفات مورفولوژیکی یا حتی رفتارهایی هستند که دیگر هیچ هدف قابل تشخیصی برای حاملان معاصر خود ( اصولات ) ندارند، مانند بقایای اسکلت پای عقبی در منقبض کننده های بوآ و نهنگ ها . در هر دو مورد، این اصول به نزول حیوانات چهار پا ( مارمولک ها یا ونگل های حتی پنجه پا ) اشاره دارد.
بقایای ژنهایی که قبلاً کار میکردند و اکنون آشکارا غیرعملکردی هستند را میتوان در ژنوم برخی از شبهزاها یافت.
جغرافیای زیستی تطبیقی
در بسیاری از موارد، توزیع گونهها سرنخهایی برای تحولات تکاملی فراهم میکند. بسیاری از گونهها دارای توزیعهای جغرافیایی هستند که نمیتوان آن را تنها با شرایط بومشناختی یا سازگاریهای محلی کنونی توضیح داد. این به ویژه در مورد بومی ها صادق است . یک نمونه شناخته شده از بومی های باقیمانده ، ، لمورها هستند تاکسونی در بین میمون های دماغ خیس . لمورها در آمریکای شمالی و اوراسیا در دوره سوم شیوع داشتند اما اکنون در ماداگاسکار بومی خشک (Haplorhini) که بعدها تکامل یافتند و آشکارا رقابتیتر هستند. آنها توسط میمونهای دماغ بودند، از همه جا بیرون رانده شدند و فقط در ماداگاسکار توانستند زنده بمانند، میمونهایی که به دلیل انزوای جغرافیایی نمیتوانستند توسط میمونهای دماغ خشک مستعمره شوند. در این بین صورت گرفت.

به همان اندازه برای زیست شناسی تکاملی، بومی های منشأ آن مهم هستند . معروف ترین نمونه در اینجا فنچ های داروین (خانواده Emberizidae) در جزایر گالاپاگوس، 965 کیلومتری سواحل اکوادور هستند. از گونههای قبلی که از طریق تشعشعات تطبیقی در مجمعالجزایری با منشأ آتشفشانی، که قدیمیترین جزیره آن پنج تا ده میلیون سال پیش تشکیل شده است، 14 گونه مختلف در سه جنس از سرزمین اصلی خارج شدهاند، توسعه یافتهاند. مهمتر از همه، شکل منقار، پر و اندازه پرندگان تغییر کرد. چارلز داروین در سال 1836 در سفر خود به دور جهان گونه های مختلف را جمع آوری کرد.
رشد جنینی مقایسه ای
کارل ارنست فون بائر اولین کسی بود که تشخیص داد مراحل اولیه رشد موجودات مرتبط بیشتر از افراد بالغ مشابه است. بر اساس این یافتهها، ارنست هکل فرمولبندی کرد که آنتوژنز تکامل ( فیلوژنی ) را خلاصه ) یک موجود زنده ( قانون اساسی بیوژنتیک میکند . با این حال، این قانون اکنون یک ساده سازی بیش از حد در نظر گرفته می شود. این مراحل بزرگسالی نیستند که تکرار می شوند، بلکه مراحل اولیه جنینی پیش شکل های مربوط به یک گونه هستند. بنابراین، رشد (هندزایی) فرد یک گونه نیز شامل مراحل قبلی موجود است.
نمونههایی از این تکرار عبارتند از ظهور روده آبشش و شکافهای آبشش در همه جنینهای مهرهدار . این ثابت میکند که تمام مهرهداران امروزی از ماهیها به وجود آمدهاند. جنین نهنگ بالین جوانه های دندانی را تشکیل می دهد در حالی که افراد بالغ بدون دندان هستند، که نشان می دهد نهنگ های بالین از نهنگ های دندان دار تکامل یافته اند.
ادراک عمومی
این واقعیت که تکامل به معنای تغییر گونه های تکاملی و در ارتباط با رویدادهای انقراض منظم رخ داده است، همیشه توسط افراد یا بخش کم و بیش بزرگی از جمعیت انکار شده است، اگرچه استدلال های علمی فقط در مرحله اولیه مطرح بود. این عمدتاً در مورد نگرش های اساسی مبتنی بر تصوری کاملاً متفاوت از جهان است که اغلب خود سیاره زمین را نیز تنها چند هزار سال قدمت می داند. گاهی اوقات جنبههای فردی، مانند یک انقراض دستهجمعی منفرد یا مکرر، که سپس با گزارشهای الهامگرفته از سیل مطابقت داده میشود، پذیرفته میشد، اما سایر اجزا و پیامدها پذیرفته نمیشد.
آنجا که اصل تکامل به رسمیت شناخته شده است، جنبه هایی از آن در حدود 150 سال گذشته، مستقل از اصول اخلاقی ، به عنوان دستور و اصل برای کنش سیاسی در جامعه بشری تفسیر و مطالبه شده است. جوامع انسانی و « نژادها تمرکز به ویژه بر مفهوم «مبارزه برای بقا» جهانی بود که » را نیز در بر می گرفت، که از ایده انتخاب نشات می گرفت و به نگرش داروینیسم اجتماعی منجر می شد . در این زمینه، ایده اصلاح نژادی که از ربع آخر قرن نوزدهم در منطقه آنگلوساکسون رایج شد و به عنوان مثال به تعیین سیاست مهاجرت کمک کرد، نیز اهمیت خاصی پیدا کرد. پس از اتخاذ تئوری ها و دیدگاه های اساسی توسط سایر دولت ها و رهبران سیاسی آنها، رادیکال ترین و مهم ترین تأثیرات ناسیونال سوسیالیسم (1933-1945) به ویژه در آلمان رخ داد. با توجه به کشتار جمعی مرتبط با آن، تکامل و نظریه مرتبط با آن تا حد زیادی نادیده گرفته شد و برای مدت طولانی، به ویژه در جهان آلمانی زبان، کمی پردازش شد. تحولات اساسی تحقیقاتی مدرن تنها برای چندین دهه در منطقه آنگلوساکسون اتفاق افتاد.
مخالفت اساسی با اصل تکامل زندگی زمینی، به ویژه تا آنجا که خود گونه یا جنس انسان ، از جمله ویژگی های آن مانند آگاهی و خلاقیت فکری، در نظر گرفته شده است، برای مدت طولانی بازنمایی شده است و برای چندین دهه به طور فزاینده ای توسط گروه های جمعیتی شکاک علمی ارائه شده است. آنها تبدیل شدن به موجودی مانند انسان را که صرفاً تابع قوانین طبیعت است یا اساساً با جهان بینی آنها ناسازگار است یا آنقدر نامحتمل می دانند که یک مقام کنترل کننده برتر را فرض می کنند، خدایی که یا گونه را آفریده یا در حداقل این فرآیند را آغاز کرده یا هدایت کرده است. این مناقشه اساسی در اروپا عمدتاً از طریق تأثیرات اختلاف بر سر تدریس نظریه تکامل در کلاس های زیست شناسی مدارس در برخی از ایالت های ایالات متحده شناخته شده است . نمایندگان مربوطه اغلب با عبارت دقیق آن بحث می کنند داستان خلقت فرموله شده همانطور که در عهد عتیق است و کلمه به کلمه آن را دنبال کنید. این شک و تردید در مورد نتایج نظریه تکامل را خلقت گرایی می نامند . شکل خاصی از آفرینش گرایی، ایده طراحی هوشمند آمریکایی است.
، کلیسای کاتولیک بارها در مورد نظریه تکامل اظهار نظر کرده است به گفته پاپ پیوس دوازدهم در بخشنامه Humani generis . پاپ ژان پل دوم در مجمع عمومی آکادمی علوم پاپی و پاپ بندیکت شانزدهم خود آمده است. در خطبه افتتاحیه خود نظریه تکامل اکنون توسط واتیکان به عنوان “سازگار با ایمان مسیحی” توصیف می شود، همچنین به تکامل خداباورانه مراجعه کنید . وین اسقف اعظم و کاردینال کریستف شونبورن با جمله “تکامل می تواند درست باشد” به نفع “ائتلاف بزرگ” زیست شناسی و الهیات صحبت کرد. درک آفرینش گرایی از کتاب مقدس، درک کلیسای کاتولیک نیست.
در اوایل اوج شکوفایی علمی اسلامی، توصیفات حیوانات و رشتههای فکری، تا حدی بر اساس مدلهای یونانی، از جمله ارسطو ، صورتبندی شد که یادآور بحثهای زیستشناختی تکاملی قرن نوزدهم، از جمله تأثیرات محیطی « لامارکی » بر تکامل، و همچنین در مورد یک “مبارزه برای هستی” که یادآور چارلز داروین است، حدس زده شده است. یکی از نمایندگان برجسته الجاحز در قرن نهم از بصره امروزی در جنوب عراق بود . در اسلام امروزی که با گرایشهای شدیداً متفاوت مشخص میشود، در مورد اینکه آیا تکامل و تا چه اندازه با دین سازگاری دارد یا خیر، نظر واحدی وجود ندارد. نگرش بدبینانه نسبت به تکامل نسبتاً گسترده است: [40] در تعدادی از کشورهای اسلامی، اکثریت زیادی از مردم نیز متقاعد شده اند که گونه امروزی انسان خردمند به شیوه ای تکاملی رشد نکرده است، بلکه ایستا است و توسط خداوند آفریده شده است . )